
审核|梅林
动物的感知、思维和反应是神经系统的功能,执行这些功能需要非常复杂的系统。人体大脑里拥有超过860亿个神经元,每个神经元又辐射出上万个突触连接,形成复杂的环路。目前的研究对大脑内的环路有一些初步的认识,但是外周神经系统——即那些从大脑、脊髓发出、广泛支配四肢、内脏与心脏的神经纤维——其解剖图谱长期以来仍停留在“大致走行”的粗略阶段,类似于一份尚未完成的草图。究其原因,身体所占据的空间远大于大脑,而外周神经的走行范围广、路径长,且常伴随难以预测的分支与交错。这使得其完整解剖图谱的绘制,在难度上不亚于、甚至超过了中枢神经系统。在信息处理层面,大脑远比外周神经系统复杂;但在物理走行和全身成像的层面,外周神经因其长程、分散、穿行于多种组织之间,反而比大脑内的神经束更难以追踪和成像(图1)。
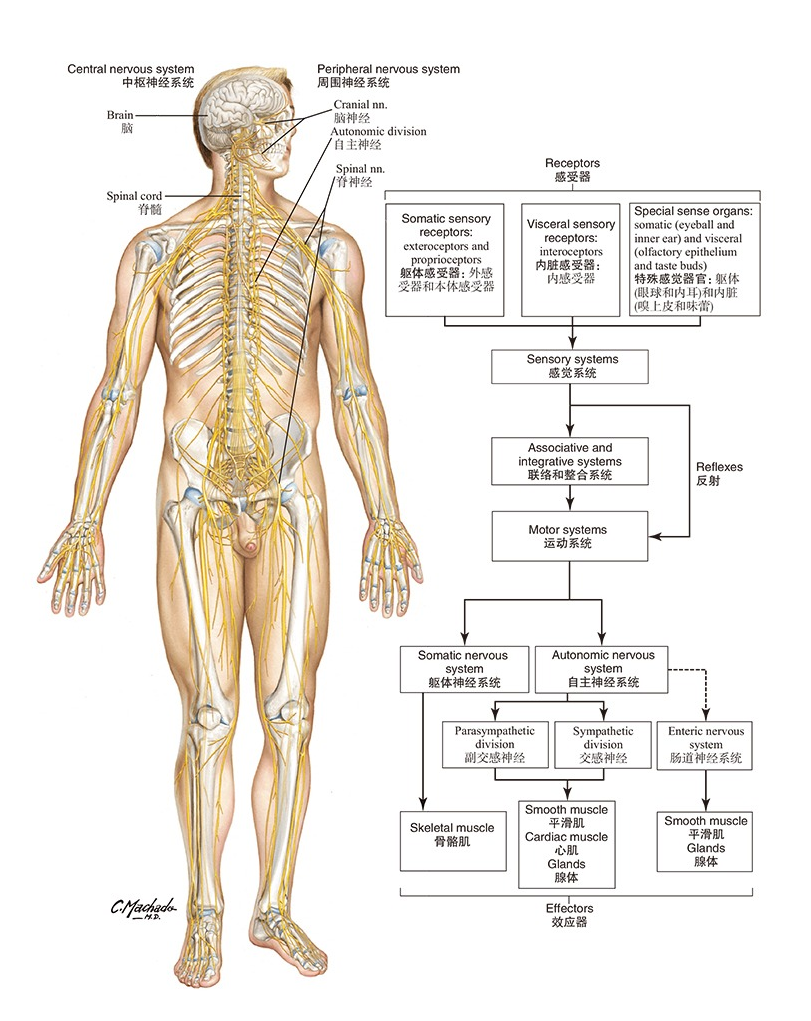
图1 脑神经和周围神经-《奈特人体解剖彩色图谱》
2026年4月8日下午,中国科学技术大学的毕国强老师在首都医学科学创新中心作学术报告,介绍他的团队自主研发、名为ARCHmap‑blockface‑VISoR的高速块面容积成像系统。该系统展现出卓越的成像性能:可在短短40小时内,以微米级的高分辨率完成对整只成年小鼠的全身成像,实现了前所未有的速度与精度的结合。基于该技术,团队对Thy1-EGFP小鼠个体脊髓神经纤维进行了精细的三维重建,揭示了感觉与运动纤维沿腹侧支与背侧支分布的独特形态特征,并完成了多条颅神经的高分辨率三维可视化;免疫染色技术实现了对全身交感神经及其分支的全面定位,清晰展示了这些神经在肢体肌肉、骨骼及大多数内脏器官中的血管周围分布模式;病毒追踪技术则揭示了迷走神经及其单个纤维的精细结构,发现了通往多种器官的意外投射路径。该技术为外周神经环路研究提供了新的结构信息,促进人们对其功能的思考和相关疾病的理解。
近期的技术进展通过采用改进的溶剂基和水基全身透明化方法,实现了对外周神经及其他身体结构的可视化观察。代表性技术包括:DISCO系列 [1](有机溶剂型)——由 Ali Ertürk 与 Hans-Ulrich Dodt 团队开发,利用四氢呋喃和二苄醚脱脂脱水,首用于小鼠脑和脊髓的透明化成像;其衍生版本 wildDISCO [2]进一步实现了基于普通 IgG 抗体的全身免疫标记。HYBRiD [3]——由 Li Ye(李烨) 团队开发,是一种有机溶剂-水凝胶重组方案,首用于新生/幼年小鼠全身及 SARS-CoV-2感染小鼠胸部的快速透明化。
在成年小鼠全身透明化成像领域,赵瑚老师团队做出了重要开创性贡献,其首创的TESOS技术(透明包埋溶剂系统技术),是一套完整的成年小鼠全身透明化成像方法,且由赵瑚老师团队首次将该技术应用于成年小鼠全身透明化成像[4]。该技术的核心基础步骤如下:1. 样本预处理:选取六周龄成年小鼠,经心脏灌注固定后完整取材,针对硬组织采用乙二胺四乙酸溶液脱钙处理,确保目标结构完整;2. 透明化处理:采用梯度溶剂洗脱法,去除组织内影响成像的杂质,实现样本初步透明;3. 透明包埋:将透明化后的样本置于双酚A乙氧基丙烯酸酯与苯甲酸苄酯组成的透明包埋溶剂中,经紫外光交联固定形态;4. 成像采集:结合改装光片显微镜与共聚焦显微镜,完成全身及局部区域成像;5. 图像重建:通过算法拼接断层图像,完成成年小鼠全身三维重建。赵瑚老师团队首次将自主发明的TESOS技术应用于成年小鼠成像,完善了成年小鼠全身透明化成像方案,为成年小鼠全身成像提供了更优化的技术路径,该技术还成功实现了成年小鼠从手指到脊髓单个感觉神经元轴突的中尺度连接组映射。尽管赵瑚老师团队的TESOS技术取得了重要突破,但当前成年小鼠全身透明化成像仍面临四大核心技术难点,具体如下:问题一,现有方法(包括TESOS技术)结合传统光片显微镜,仅能粗略呈现外周神经的整体架构,无法均匀分辨纤细神经分支和单根轴突纤维等精细结构;问题二,共聚焦显微镜及块面铣削技术存在固有成像速度局限,难以满足成年小鼠全身大体积样本的高分辨率成像需求;问题三,大体积样本深层成像需依赖长工作距离镜头,该镜头会导致分辨率下降,且组织异质性引发的折射率差异,会产生随成像深度递增的光学像差,影响成像质量;问题四,小鼠身体具有高度异质性,胸腔、腹腔等天然空腔的存在,易导致透明化后的样本发生形变,影响成像准确性。
综上,需要解决的核心问题包括:分辨率不足、成像速度慢、深层光学像差以及切片形变。毕国强老师的团队正是针对这四大难题,开发了 ARCHmap-blockface-VISoR 整合方案。
为了解决这些难题,团队打出了一套漂亮的“组合拳”。第一拳名为 ARCHmap(Active CleaRing with CUBIC-LH, Hydrogel-embedding, and RI-matching with PuClear),是一套水性透明化流程。首先,他们对CUBIC-L(10% w/v曲拉通X-100 +10% w/v N-丁基二乙醇胺)进行改良—CUBIC-L是一种亲水型组织透明试剂,可实现高效脱脂、脱色与组织通透,提升光学透明性。团队将 N-丁基二乙醇胺浓度从10% w/v提高到 15% w/v,得到CUBIC-LH,在实现优异组织透明化的同时保留荧光信号,并将组织尺寸变化控制在最小。与传统的低温被动固定不同,团队采用连续主动心脏灌注的方式,将固定液、脱钙液及 CUBIC-LH 透明化试剂直接泵入小鼠血管系统,使其流遍全身。这种主动灌注方式显著提高了试剂的渗透均匀性,尤其是在大脑和脊髓等深部组织中,透明效果和荧光保留都更为出色。第二,在组织透明化之后,团队用自行研发的水凝胶包埋溶液(含水凝胶单体、牛血清白蛋白及碘海醇)对整只小鼠进行包埋。这一步相当于给软塌塌的身体穿上一件“紧身衣”,极大地提高了透明化小鼠身体的机械强度,从而有效防止了后续连续切片过程中的组织变形。最后,将包埋好的样本浸泡在团队自研的 PuClear 折射率匹配液(50% w/v碘海醇 + 23% w/v尿素 + 11% w/v三乙醇胺 + 16% w/v水)中,使组织与介质的折射率达到一致(RI=1.52),光线在组织内部几乎不发生散射。经过ARCHmap处理的整只小鼠身体光学透明均匀,能够清晰观察到体表以下至少 600 微米深处的精细结构。
透明之后怎么拍?这就是第二拳:blockface-VISoR(blockface-Volumetric Imaging with Synchronized On-the-fly Scan and Readout)成像系统。小鼠被固定在一个精密试管里,每拍完一层,上方的振动切片机就切掉400微米厚的顶层,露出一个新鲜平整的“块面”。然后,显微镜用两枚斜对的物镜和高速相机,以200帧/秒的速度扫描这个新鲜切面下方600微米深的三维体积。样品在扫描过程中以0.7毫米/秒的速度连续移动,没有任何停顿,自然也没有运动模糊。扫描完成后,样品自动退回切片区,再切下一层,如此循环,每次成像与下一次成像之间有200微米的重叠,确保后续三维拼接时不会出现裂缝。整个流程全自动,一个完整的小鼠身体可以在约40小时内完成透明化成像(约200个循环),每个荧光通道生成约70 TB的数据。用传统方法拍同样的体积,可能需要几个月,然而40小时绝对已经是这个尺度下的速度天花板(图2、3)。
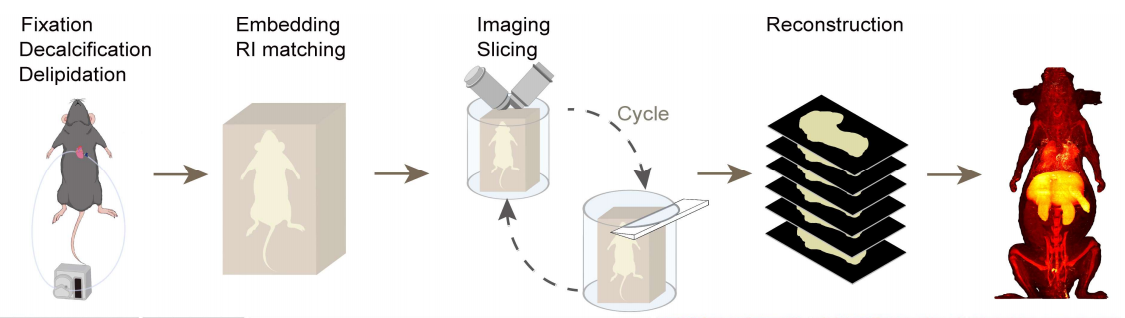
图2 用于全小鼠成像的 ARCHmap-blockface-VISoR 流程图
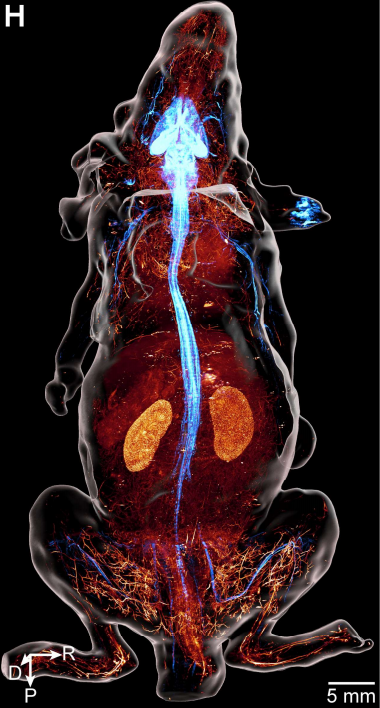
图3 成年Thy1- EGFP 小鼠神经与血管系统的三维视图
颅神经是直接从脑和脑干发出、共 12 对的外周神经,主要负责支配头面部的感觉、运动以及部分内脏活动,是连接大脑与头颈部器官的关键传导通路,参与视觉、听觉、面部运动、咀嚼、味觉、牙周感觉等重要生理功能。有了这套“天眼”,他们首先拿Thy1-EGFP转基因小鼠开刀。这种小鼠的神经元会稀疏地表达绿色荧光蛋白,正好适合观察单个神经纤维。在头部,他们清晰地看到了滑车神经、面神经和三叉神经的单根纤维。三叉神经的运动部分粗壮、分支密集,通过“爪样”的神经肌肉接头抓住面部的咀嚼肌。这本身不令人意外,但真正让他们兴奋的是三叉神经的感觉部分——他们发现有一束感觉纤维,竟然直接跑到了门牙和磨牙的牙周组织里,而且追踪单根纤维后发现,这条感觉路径完全独立于运动纤维,走的是另一条路(图4)。这或许提示,牙周的感觉纤维可能并不与三叉神经的运动纤维伴行,而是拥有自己独立的专属传导路径。
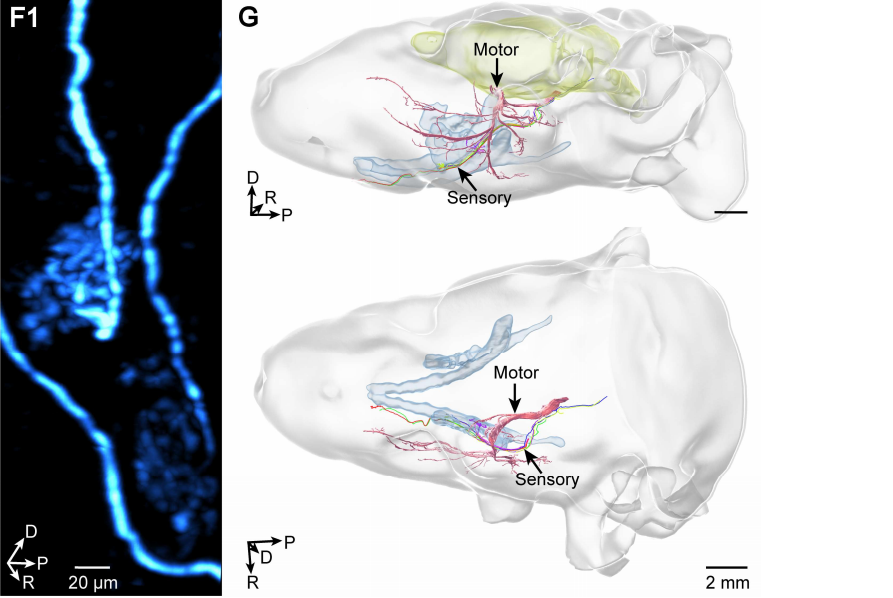 |
|
| 图4-1 | 图4-2 |
图4 4-1三叉神经向牙周组织的感觉传入纤维;4-2支配牙周组织的三叉神经感觉纤维的单纤维追踪
脊髓神经是连接脊髓与躯干、四肢的外周神经,共分颈、胸、腰、骶、尾多个节段,T2–T13 即代表胸段第 2 至第 13 对脊神经,主要负责胸壁、腹壁及背部的感觉与运动控制。脊髓神经按功能可分为两大类:感觉神经负责将皮肤、肌肉、关节的信号传入脊髓,运动神经负责将中枢指令传出至肌肉;而调节内脏、血管、腺体活动的则是交感神经与副交感神经(属于自主神经系统,不随脊神经分节段走行)。每根脊髓神经发出后会分成腹侧支与背侧支:腹侧支较粗大,支配躯干腹侧与四肢肌肉与皮肤;背侧支较细小,主要支配背部深层肌肉与皮肤。接着,他们从两只Thy1-EGFP小鼠的T2到T13节段,手动重建了191个脊髓神经元的完整外周投射,其中包括腹侧支的66个运动神经元和66个感觉神经元,以及背侧支的30个运动神经元和29个感觉神经元(图5)。这相当于给每个神经元建立了一份“个人档案”:它的主纤维有多长,分支有多少,往哪个方向走,在哪个节段转弯。
定量分析给出了几个很有意思的规律。在腹侧支,运动纤维的主纤维长度从T2到T13呈节段依赖性增加,而感觉纤维的长度在各节段间相对均匀。而在背侧支的运动神经纤维表现出广泛的分支,而且从T2到T10,分支数和总分支长度都在显著增加。背部的肌肉精细控制,看来确实需要更复杂的“接线图”。
更有趣的是形态上的差异。超过一半的腹侧支感觉神经会走一个“大V字形”急转弯,好像在故意绕路;而在背侧支,感觉神经的轴突呈现出“多锯齿状”的曲折形态,像心电图一样来回折,而运动神经则短粗、分支密集,完全不是同一个风格(图6、7)。这些形态上的多样性,是传统的肋间神经或肋下神经解剖学描述无法捕捉到这一特征。
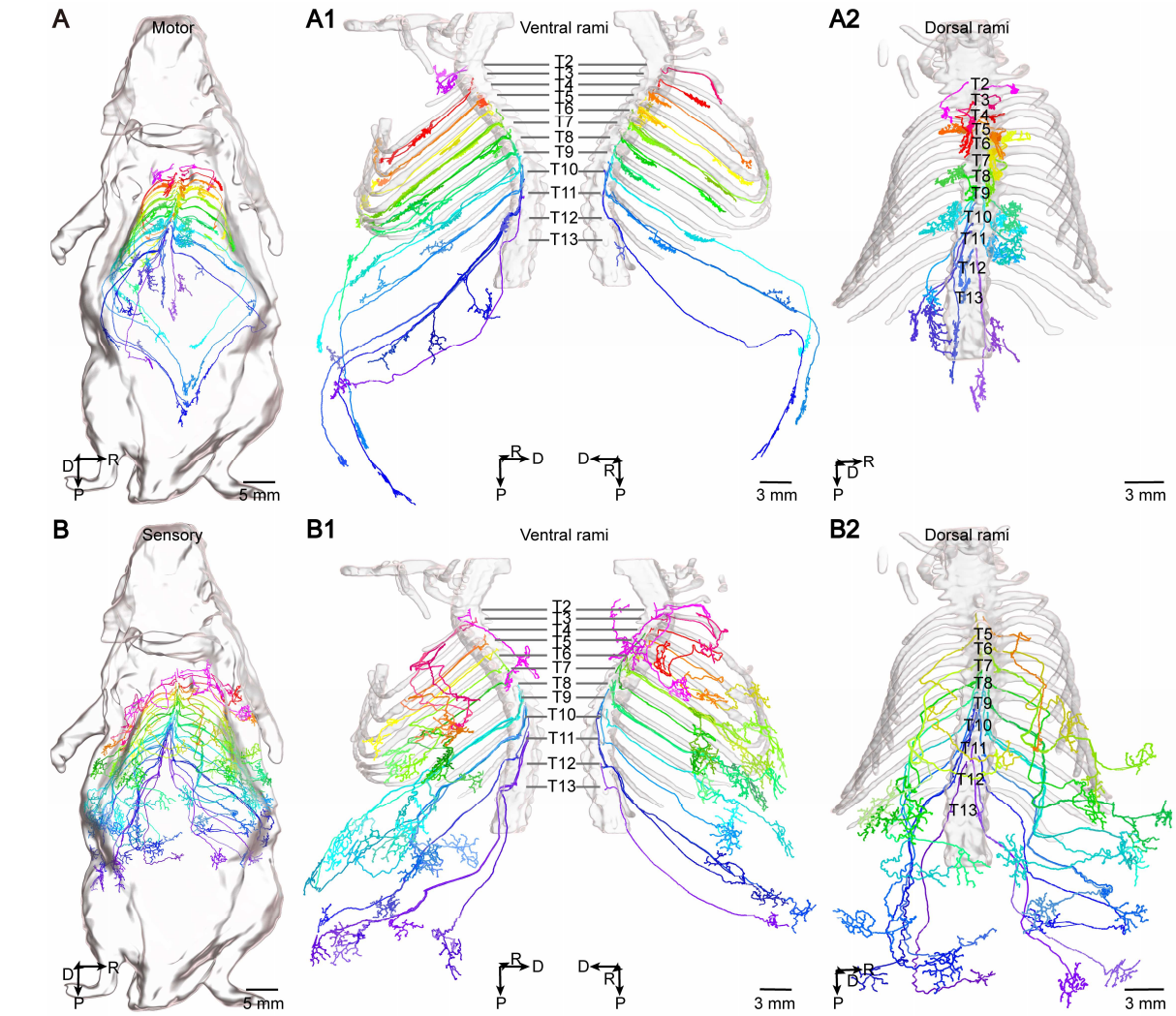
图5 对2只Thy1- EGFP 小鼠通过胸椎骨骼进行定位后,对其胸段脊髓运动神经元(A)和感觉神经元(B)进行了全身追踪研究

图6 腹侧脊髓感觉神经元的代表性解剖形态学特征
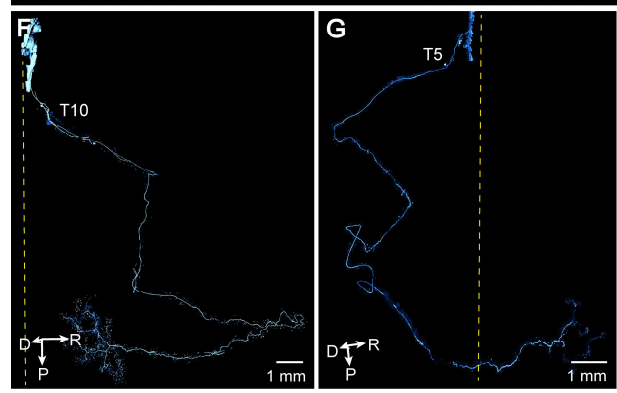
图7 背侧脊髓感觉神经元的代表性解剖形态学特征
如果说Thy1-EGFP小鼠展示的是稀疏标记下的单纤维之美,那么全身免疫染色则展示了另一种壮阔。他们用抗酪氨酸羟化酶(TH)抗体标记了交感神经,同时用凝集素标记了全身血管(图8)。结果清晰显示:在小鼠的骶骨、骶髂关节、腿部肌肉、肾脏皮质、脾脏白髓、肝脏和胰腺中,交感神经纤维几乎都紧密缠绕在血管周围,形成血管周围的网状结构。这种血管伴随模式在躯干和四肢的各个区域均被一致观察到。古典解剖学早已认识到交感神经在动脉周围形成网状结构,但此前从未有人以亚细胞分辨率和全身尺度系统验证其普遍性。本研究的观察表明,血管周围网状交感神经纤维广泛分布于头颈、躯干、四肢及大多数内脏器官,唯一的例外是胃和肠道——该处的交感纤维主要沿胃肠壁形成网状结构,与血管基本不重叠。这种交感神经结构的中尺度图谱对浅表神经刺激和射频消融等临床操作具有重要价值,这些技术常用于治疗疼痛和高血压等疾病。
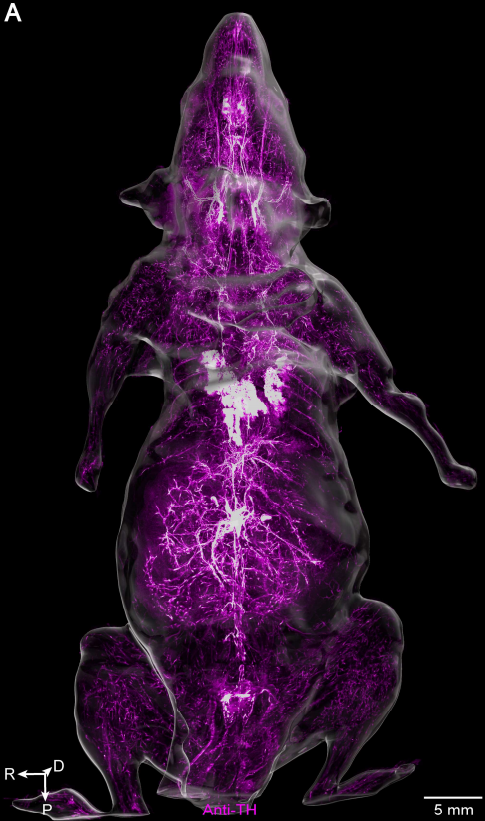
图8 通过抗TH抗体进行全身免疫染色获得的成年小鼠交感神经系统三维视图
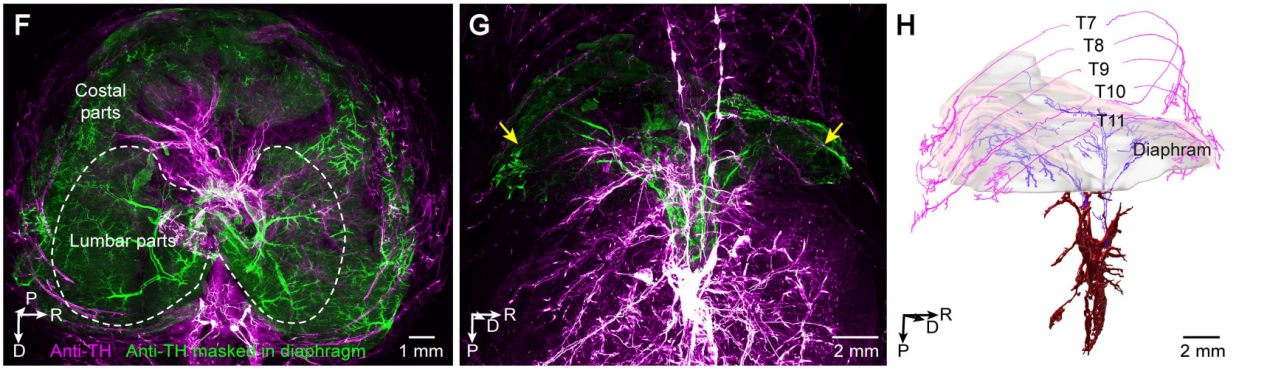
更精彩的是膈肌。膈肌是我们呼吸的主要肌肉,以往大家知道它接受膈神经的运动支配,还接受交感神经支配。然而,这些交感神经支配的来源及其区域特异性尚不明确。这篇研究第一次清晰地区分出:膈肌腰部的交感神经来自腹腔-肠系膜上神经节(CSMG)和膈神经节,而肋部的交感神经则来自胸椎旁链(图9)。
 |
||
| 图9-1 | 图9-2 | 图9-3 |
图9-1 膈肌交感神经支配的三维视图(膈肌标注绿色);9-2标示从胸椎旁链节神经节至肋膈肌的交感神经传入点;9-3图示 CSMG(紫色)与胸椎旁神经节(品红色)对膈肌交感神经支配的纤维束走行轨迹
最后,研究团队用AAV病毒(adeno-associated virus)分别标记了迷走神经的运动核(DMV)和感觉神经节(NG/JG),追踪了迷走神经从颈部到腹部的完整路径(图10)。在颈部,运动纤维和感觉纤维保持空间分离;而在胸部远端节段,两者逐渐混杂在一起。他们还发现,左右喉返神经之间存在多条交叉纤维,同时左右迷走神经干之间也存在相互连接。通过EYFP标记左侧迷走感觉神经元,研究团队在检查的三只小鼠中的两只中,观察到了从左侧喉返神经发出的纤维在不同位置交叉到右侧,支配喉部。这些交叉纤维的特征在此前未被识别。
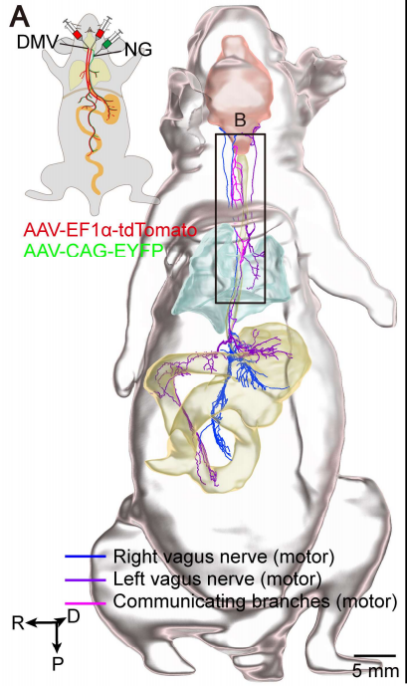
图10 迷走神经致密标记内脏运动纤维的全身追踪研究
在单纤维示踪方面,研究团队采用AAV-retro技术稀疏标记了胃投射的迷走神经元,成功追踪到了三条从迷走神经背核(DMV)到胃的完整纤维(图11)。这三条纤维在迷走神经干内均无分支。其中两条沿同侧迷走神经干行进,分别支配胃的腹侧和背侧。腹侧与背侧的分离支配,提示迷走神经对胃的调控可能存在空间上的功能分区。第三条纤维起自左侧迷走神经背核(DMV),在膈上位置交叉到对侧,随后沿右侧迷走神经纤维附近向头侧方向行进一段后急转,再向尾侧返回,最终终止于胃背侧壁。这些发现表明,内脏神经的布线路径比以往认知更为复杂。
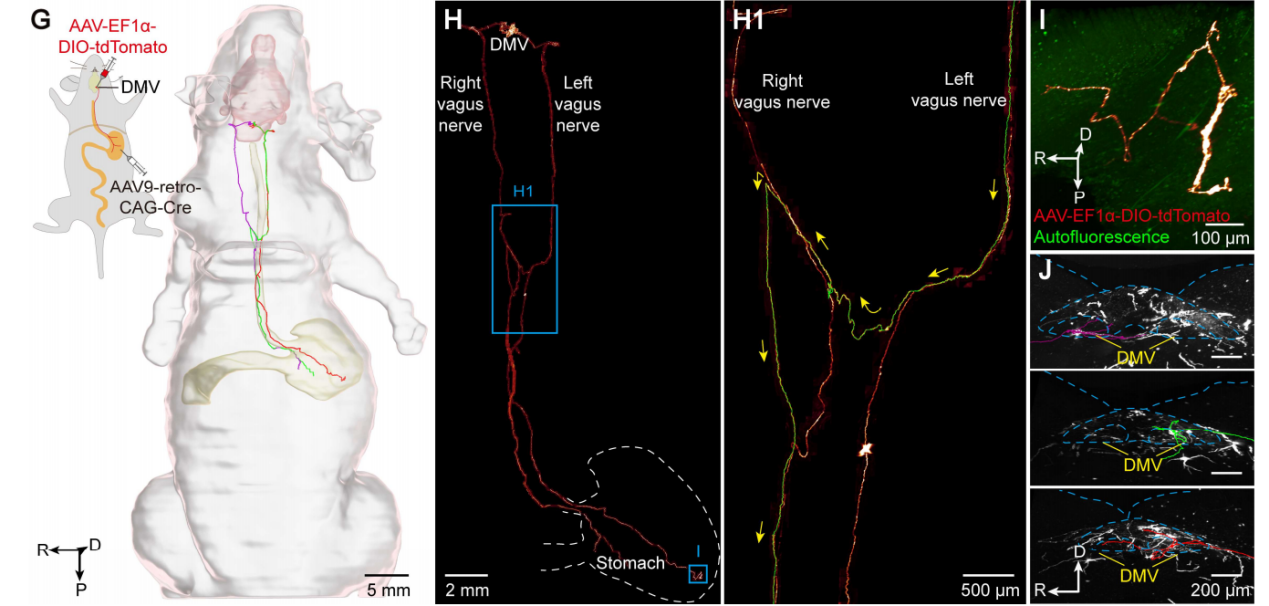
图11 对三条投射至胃的迷走神经在整个体内的单个神经元追踪
过去十年,脑连接组学让我们得以窥见思维的中枢舞台;而外周神经网络——那些连接每一寸肌肉、每一处内脏、每一片皮肤的“生命线”——却长期隐匿于视野之外。如今,这支研究团队用ARCHmap-blockface-VISoR系统,在40小时内为成年小鼠的全身神经拍下了第一张亚细胞分辨率的“全家福”。从此,我们不仅能看见单根纤维如何穿行于骨骼与肌肉之间,也能追踪它在何处转弯、在何处分支、在何处与血管相伴或分道扬镳。
这是一种从“大概走向”到“精确地图”的跃迁。研究团队已用这套系统完成了52只小鼠的成像,并公开了在线浏览平台,让全球科学家都能在这张新地图上寻找自己的答案。更重要的是,这套系统可以扩展至大鼠甚至猕猴,意味着未来我们可以用同样的精度去探索更大、更接近人类的神经系统。
外周神经系统中,交感神经纤维与迷走神经纤维分别沿着特定的、进化上保守的路径分布。今天,我们首次实现了对这些纤维完整形态的高分辨率可视化—这标志着外周神经连接组学研究的一个新起点,而非终点。
这套系统还差点什么
当然,没有任何技术体系是完美的,该成像体系也存在几点固有局限。由于VISoR采用斜向成像配置,物镜的数值孔径(NA)会受到几何限制,无法使用最高NA镜头,导致横向分辨率存在上限;轴向分辨率则取决于照明光斑厚度,通常为几个微米,若需观察突触、蛋白精细定位等更细微结构,需将目标区域切取后,借助共聚焦或超分辨显微镜进一步拍摄。此外,少数深部脂肪组织偶尔会出现成像模糊现象,主要源于脱脂不够彻底,需在未来进一步优化透明化流程。同时,样品经水凝胶包埋后,抗原位点会被锁定,难以通过新抗体对切片进行二次染色,实验前需提前规划好所有待染色标记,无法后续补充染色。
本文图1来源于《奈特人体解剖彩色图谱》,其余图片均来源于毕国强老师团队2025年发表的论文[5]。


 北京市丰台区右安门外西头条10号
北京市丰台区右安门外西头条10号
 010-86738999
010-86738999
 cimr@cimrbj.ac.cn
cimr@cimrbj.ac.cn









 En
En
