
撰文|范炎炎(2024级博士研究生)、宋明坤(2025级博士研究生)
审核|张昱
2026年4月15日下午,应首都医学科学创新中心(CIMR)张昱老师的邀请,来自美国哈佛医学院、波士顿儿童医院的Frederick W. Alt教授来访,并作了题为“Chromatin Loop Extrusion Differentially Generates Diverse IgH, Igκ, and Receptor Edited Repertoires by Distinct Mechanisms”的学术报告。Alt教授是免疫学和分子遗传学领域的国际顶尖学者,长期深耕于V(D)J重排、DNA损伤修复机制以及染色质三维结构等研究领域。近年来,他的团队深入揭示了RAG核酸内切酶如何利用染色质环挤压(Loop extrusion)机制在B细胞发育过程中介导抗体基因(如IgH和Igκ)的重排。这些开拓性的研究深化了我们对免疫系统多样性产生机制以及基因组稳定性维护的认知,为相关免疫系统疾病及肿瘤的发生机制提供了重要的理论基础。

图1:Frederick W. Alt教授报告现场
摘要
V(D)J重排通过在跨越数兆碱基(Mb)的抗体重链(IgH)和轻链(Igκ)基因座中组装大量V、D、J基因片段,构建抗体可变区外显子,是适应性免疫多样性的核心来源。该过程由RAG核酸内切酶介导,其识别位于各基因片段两侧的重组信号序列(RSS),并遵循“12/23规则”实现精确切割。RAG如何在大尺度基因座上远距离定位RSS的机制长期未被阐明。Frederick W. Alt教授重点阐述了团队的核心发现:RAG并非随机扩散寻找目标,而是依赖于cohesin(黏连蛋白)介导的染色质“环挤压”机制进行线性扫描,从而高效搜索并捕获目标RSS。这一机制在IgH和Igκ基因座中均得到验证。
在IgH基因座中,RAG首先锚定于以JH-RSS为核心的重组中心(recombination center, RC),随后沿环挤压递呈的染色质对上游D和VH片段进行线性扫描,并优先识别与J-RSS呈相对(删除)方向的RSS,从而实现有序的V(D)J重排。
在Igκ基因座中,初级重排主要由Vκ与Jκ1连接启动,此过程中,cohesin介导的环挤压将不同取向的Vκ片段递送至以Jκ1为核心的RC附近,但上游Cer/Sis调控元件会物理性阻断RAG向远端Vκ区域的线性扫描,使其形成一个暂时的RC,通过扩散作用完成首次重排;当初级Vκ–Jκ1重排失败时,系统会启动次级重排(受体编辑)。此时,初级重排引发的Cer/Sis删除或位移作为关键“发育开关”,将重排模式从基于“双环”的扩散机制转变为基于“单环”的线性扫描机制。新的重排中心可在Jκ2、Jκ4或Jκ5处建立,使RAG能够绕过原有结构屏障,对上游Vκ片段进行大范围线性扫描。这一发现不仅阐明了RAG在大尺度基因座上的靶向搜索机制,也为理解抗体多样性产生及中枢免疫耐受(受体编辑)的分子基础提供了机制层面的新见解。
精彩回顾
一、RAG核酸内切酶介导的IgH V(D)J重组中心12/23 RSSs精准切割机制
报告首先回顾了B细胞发育早期发生V(D)J重排的基本原理与时空调控机制。在骨髓的Pro-B细胞阶段,抗体IgH基因必须严格遵循“先D-to-JH,后VH-to-DJH”的重组顺序(图2c)。为了确保这一时序逻辑,基因组三维结构发挥了重要的物理隔离作用:IGCR1(Intergenic Control Region 1)元件与3' Igh CTCF(CCCTC-binding factor)位点锚定形成了一个封闭的3'染色质环结构域(图2e)。该拓扑结构将D片段“圈禁”在以JH和iEμ增强子为核心的V(D)J重组中心(RC)内,同时将上游的VH片段有效隔离在外,防止其过早干扰重组(图2a)。在这种受控的三维空间下,RAG精准锚定于RC内部,将其作为执行切割的物理平台。RAG能够特异性识别抗体基因片段两侧的重组信号(RSS)。RSS由较为保守的七聚体序列(heptamer)和九聚体序列(nonamer)组成,中间间隔12 bp或23 bp。在该重组中心内,RAG利用其独特的Y型异二聚体结构(图2d),严格遵循经典的“12/23规则”。它首先在RC内捕获带有一个23-RSS的片段(如JH),随后通过cohesin介导的环挤压机制,接收由线性扫描递呈而来的互补12-RSS片段(如D片段)。RAG必须将12-RSS与23-RSS进行如同“钥匙与锁”般的精确配对(图2b)。通过这种精密的3D空间对齐与生化突触复合物的形成,RAG在RC内部诱导特异性的DNA双链断裂,从根本上防止了错误重排,确保了抗体IgH基因片段能够高度有序且准确地完成组装(图2)。
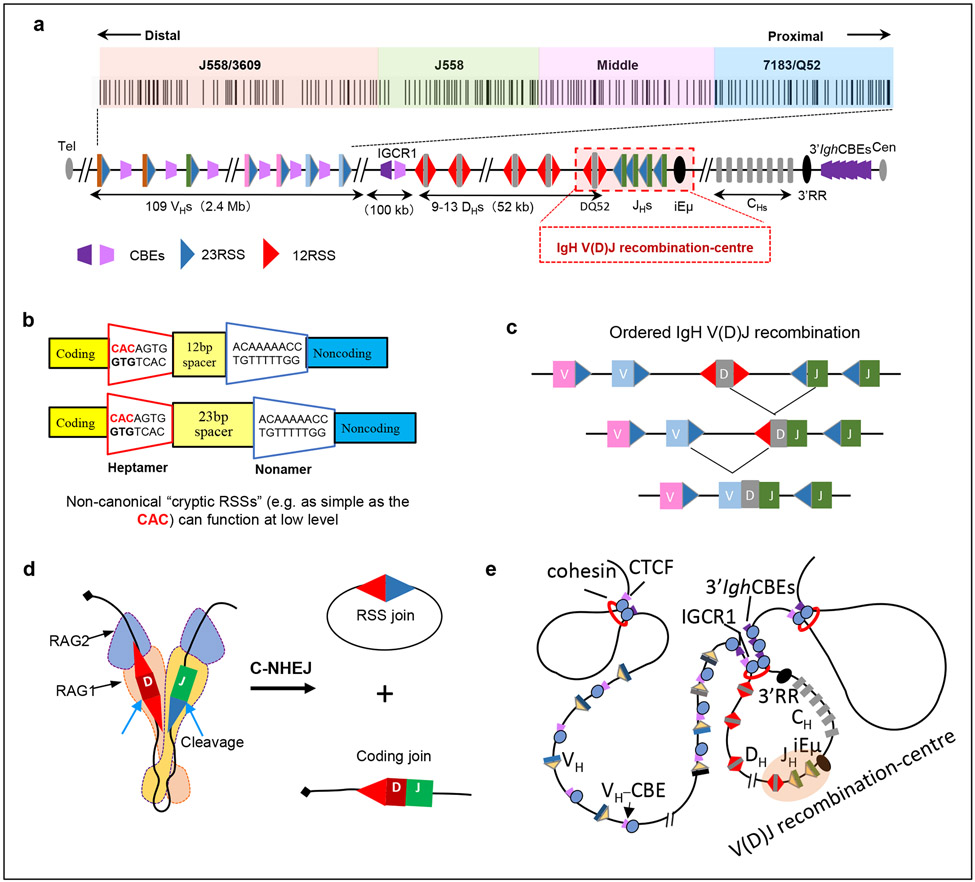
图2:V(D)J重排的基本规则:“12/23规则”与重组信号序列
(Zhang, Y., Zhang, XF., Dai, HQ., et al. Nature Reviews in lmmunology, 2022)
二、环挤压介导IgH的线性扫描与有序重组
IgH基因的精准组装是由生物物理动力学与生化反应精细调控的有序过程。在重组初期,IGCR1与3' Igh CTCF位点形成封闭的3'环结构域,将D片段限制在RC内,并将上游VH片段隔离在外,防止其过早参与重组(图3e)。在此空间内,由cohesin介导的环挤压像传送带一样拉动染色质,这种线性扫描机制通过对“相对方向”RSS的严格识别,发生删除型重排(图3a)。然而,当环挤压将特定片段拉至RC的极近距离时,DNA会进入RAG的“扩散半径”(图3b),从而引发了不依赖扫描、由短程扩散介导的低频倒位重排(图3b)。当IGCR1屏障的限制消除后,RAG的扫描活性得以向更远端的VH区域延伸(图3c);在此区域中,CTCF结合位点作为“物理减速带”发挥了决定性作用,它通过阻碍环挤压、增加特定VH片段在RC停留的时间,直接赋予了该基因优先重组的能力(图3c)。正是这种由“环挤压引擎”、“CTCF 减速带”以及“RAG扫描仪”构成的物理反馈系统,因果相循地保障了抗体基因重排在Mb级尺度上的精准性、顺序性与多样性。
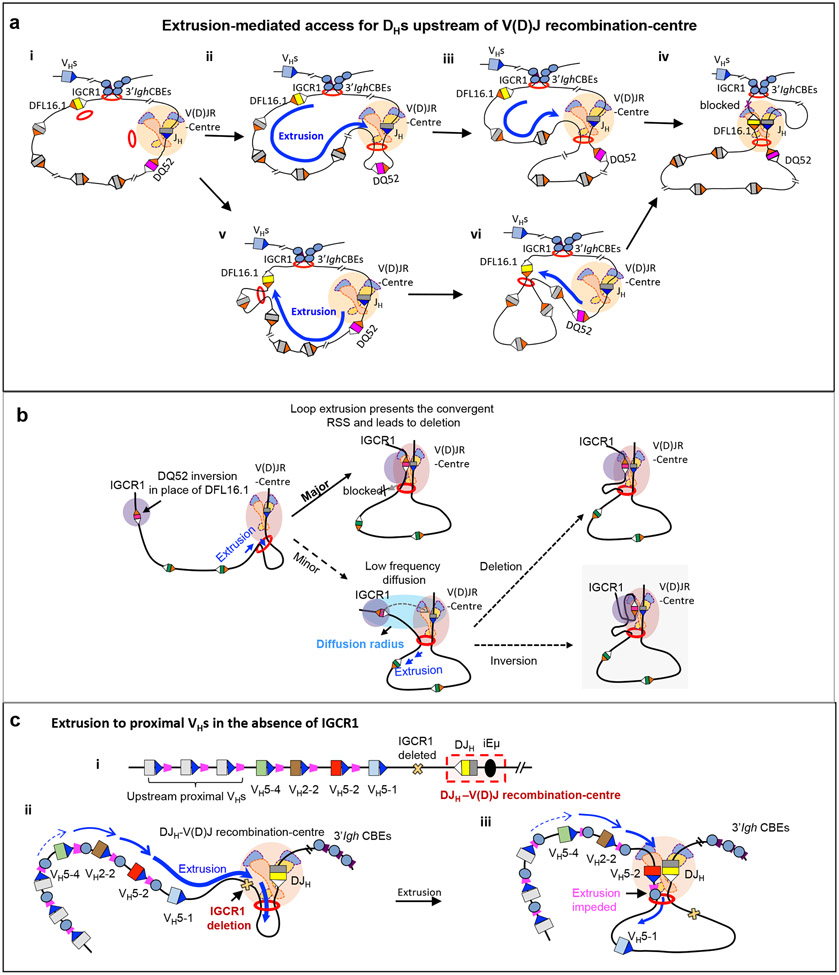
图3. 环挤压介导的RAG扫描确保IgH基因重排的精准性、顺序性与多样性
(Zhang, Y., Zhang, XF., Dai, HQ., et al. Nature Reviews in lmmunology, 2022)
三、Igκ基因的初级重排:依赖于Cer/Sis CBE平台的扩散机制
与IgH基因座不同,Igκ基因座虽然仅包含4个J片段,但其Vκ基因库极为丰富,Igκ的初级重排(Vκ与Jκ1的连接)依赖于基于Cer/Sis CBE平台的双环扩散机制,这使得广泛分布的Vκ都能被呈递给Jκ1重排中心。此过程中,Vκ与Jκ1的连接可能有两种方式:当两者呈相对方向时,发生删除型连接;呈相同方向时,发生翻转型连接。Alt教授的研究阐明,初级的删除型或翻转型Vκ-Jκ1重排会在物理上删除或移位Cer/Sis元件。这一拓扑学改变如同一个“发育开关”,将原本依赖双环扩散的初级重排机制,直接转换为类似IgH的单环线性扫描机制,以用于后续的次级受体编辑重排(连接至Jκ2-5)。
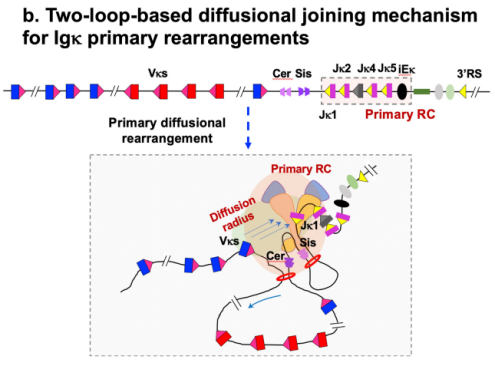
图4:Igκ初级重排机制的拓扑学转换
(Li, X., Hu, H., Zhang, Y. et al. Nature, 2026)
四、次级重排:线性RAG扫描限制了Igκ次级重排的Vκ库
在次级重排过程中,新生成的基于Vκ-Jκ1的次级重组中心(RC)会沿着上游染色质进行单向线性扫描。然而,次级重排并没有随机利用整个基因座,而是高度局限于紧邻重组中心上游的有限Vκ片段。机制层面的深入探究发现,高度活跃的Vκ转录会形成物理扫描障碍,增加了这些区域的染色质可及性与RC的互作频率。同时,Igκ本身具有强效的重排信号序列,使得RAG在扫描到第一个合适的Vκ时便迅速达到饱和重排,进一步解释了受体编辑为何主要发生在邻近片段上。
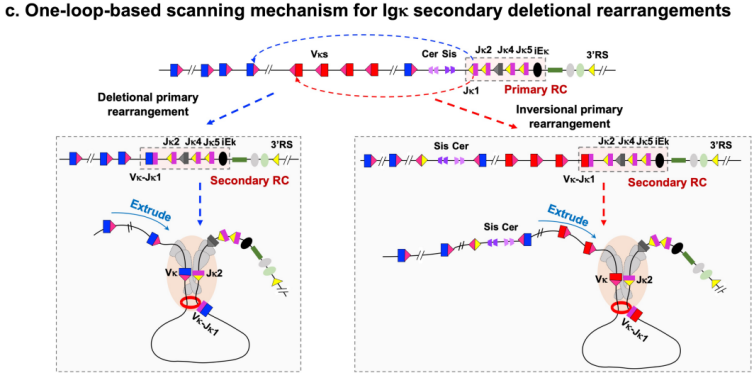
图5:Igκ次级重排机制的拓扑学转换
(Li, X., Hu, H., Zhang, Y. et al. Nature, 2026)
讲座结束后,在场师生围绕IgH、Igκ受体编辑机制与Alt教授展开了进一步讨论,Alt教授对每个人的问题都进行了细致、深入的解答,逻辑清晰,深化了听众对于V(D)J重排过程精密调控机制的理解。
参考文献
1. Zhang, Y., Zhang, XF., Dai , HQ., et al. Nature Reviews in lmmunology, 2022; 22(9): 550-566. doi: 10.1038/s41577-022-00679-3.
2. Li, X., Hu, H., Zhang, Y. et al. Nature, 2026; 653(8115): 870-878. doi:10.1038/s41586-026-10362-5


 北京市丰台区右安门外西头条10号
北京市丰台区右安门外西头条10号
 010-86738999
010-86738999
 cimr@cimrbj.ac.cn
cimr@cimrbj.ac.cn









 En
En
